- Phylum
- Dinoflagellata
- Class
- Dinophyceae
- Order
- Peridiniales
- Habitat
- plankton, pelagial
- Distinctive features
- Co-occurs with P. cunningtonii
- Organization
- flagellated single cells
- Color
- yellow to golden-brown
- Cell shape
- prolate spheroid
- Cell diameter (D)
- 21 - 30 μm, median: 26 μm (N=>2000)
- Cell length (L)
- 28 – 37 μm, median: 32.5 μm (N=700)
- Cell biovolume
- 6600 - 17500 μm3, median: 11500 μm3.
- Biovolume equation
- V, μm3 = 1.2514 D2.793 (regression eqn. based on >500 measurements of both D and L with V computed for a prolate sphaeroid).
Morphological features
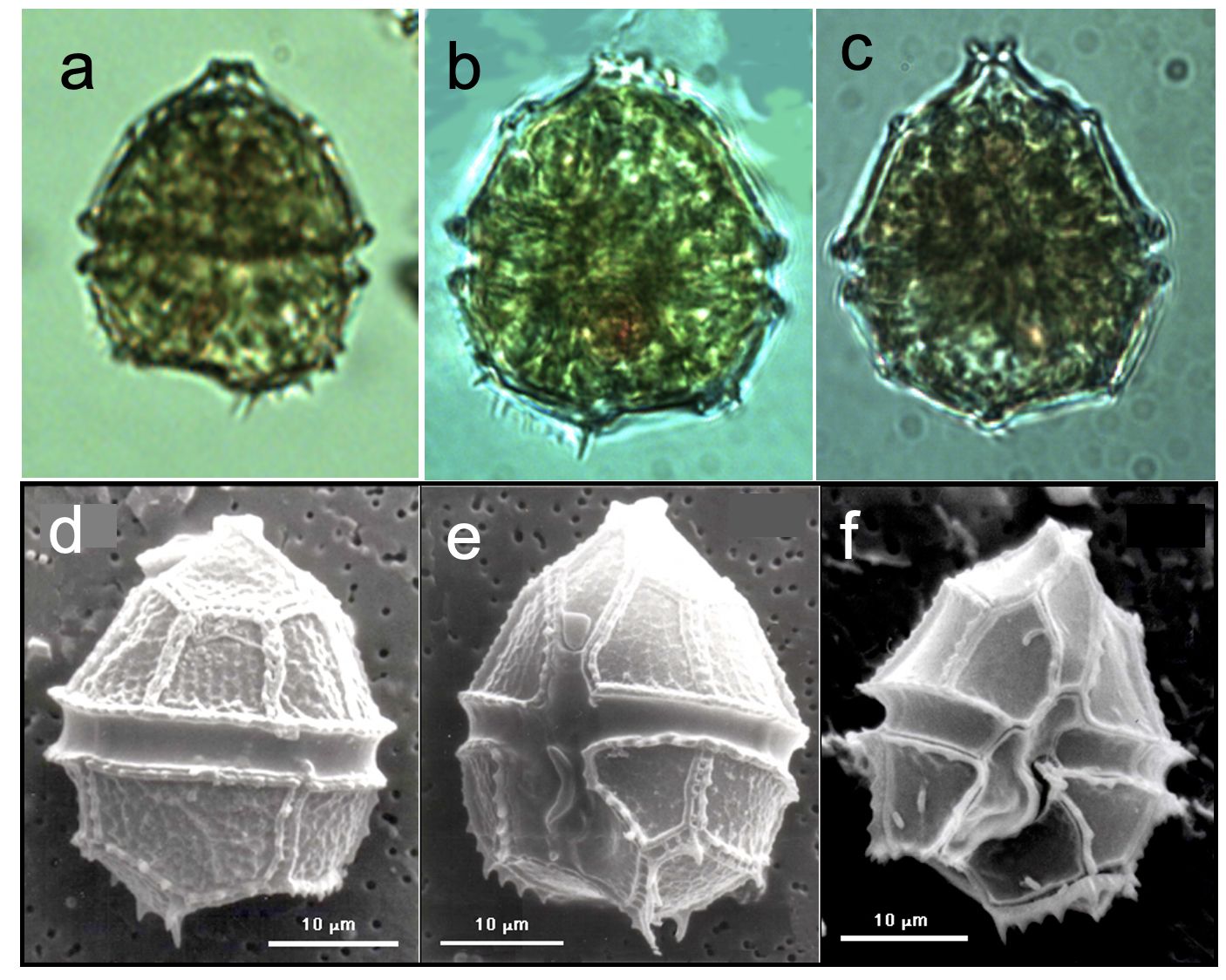
Cells are (having 5 sides) or ovoid in shape and slightly flattened dorsi-ventrally. The is conical, with a “mouth” shaped (uppermost end) and is larger than the , which is rounded but excavated on the (base, opposite to the apex). The encircles the cell; the widens along the hypotheca. Several spines or teeth-like projections are present on the hypotheca, typically some of them are lined up as in a brush (Plate 1). The chloroplasts are numerous and discoid. A red body is occasionally seen (Plate 1). SEM shows that the plates are concave and ornamented by rows of fine punctae, and that the sulcus extends all the way to the concave base of the hypotheca. The resting cysts (not shown) are somewhat oval with a thick cell wall, 36 μm x 28 μm. Parvodinium elpatiewskyi is similar to P. cunningtonii in size and shape, and they tend to co-occur. See here our tips for distinguishing between these two species under the light microscope.
Ecology
Parvodinium elpatiewskyi is one of the common algal species in the Kinneret plankton, present in the water at all times of the year. In the absence of Peridinium gatunense in recent years, Parvodinium elpatiewskyi seems to be more abundant than in Peridinium bloom years, although 1999 was an exception (Fig. 1). It always occurs with a second species of the same genus - Parvodinium cunningtonii (Figs. 1, 2). Its abundance has a typical annual cycle with a peak between April and June (Fig. 3). Cell size also fluctuates seasonally, being smaller in summer, larger in winter (Fig. 3). Peak cell size (February-March) precedes peak abundance by 2-3 months. These seasonal cycles are shared with Parvodinium cunningtonii and Peridiniopsis borgei. The multiannual average contribution of this dinoflagellate to total phytoplankton biomass is 3%. Its maximum biomass ever recorded was 53 g m-2 - 41% of total biomass, in April 2006, a “no-Peridinium” year.
Environmental conditions
Parvodinium elpatiewskyi > 5 g m-2 or > 25 cells mL-1 occurred more frequently but not exclusively at calcium concentrations > 45 mg L-1, pH > 8.5, dissolved oxygen > 10 mg L-1, showing greater abundance at low NH₄ and TDP, wide range of DON (Fig. 4). Furthermore (not shown), the higher biomasses usually occur at chloride > 230 mg L-1, > 1050 μS cm-1, > 115 mg CaCO3 L-1, at a relatively narrow range of long wave radiation (300-370 Watt m-2), but over the full range of short-wave radiation (100-350 Watt m-2) and water temperatures (14-30oC), when the lake is stratified. Cell size declines with increasing temperature, especially at temperatures below 23C, and increases with calcium and alkalinity (Fig. 5).
Additional figures
Cite this record as: Dr. Tamar Zohary, Dr. Alla Alster. 16 June 2026. Electronic publication. Israel Oceanographic & Limnological Research. https://kinneret-algae-atlas.org/ Searched on —.