- Phylum
- Dinoflagellata
- Class
- Dinophyceae
- Order
- Peridiniales
- Habitat
- plankton, pelagial
- Distinctive features
- Co-occurs with Parvodinium elpatiewskyi
- Organization
- flagellated single cells
- Color
- golden-brown
- Cell shape
- prolate spheroid
- Cell diameter (D)
- varies significantly over an annual cycle 21 - 30 μm, median: 25.6 μm (N=>2000)
- Cell length (L)
- 27 – 36 μm, median: 31 μm (N=700)
- Cell biovolume
- 6300 - 17500 μm3, median: 10500 μm3.
- Biovolume equation
- V, μm3 = 2.099 D2.623
Morphological features
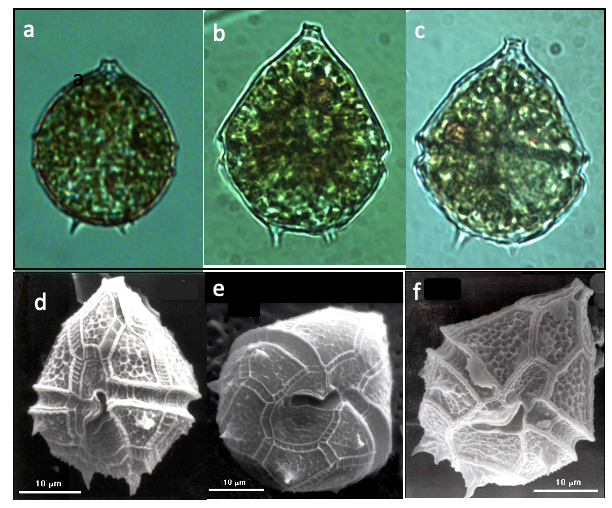
Cells shape is of half sphere () + half cone () of about the same diameter above it, with the traversing the cell where they connect (Plate 1). The of the epitheca has a “mouth” shape. Two to six spines extend from the hypotheca. The barely extends into the epitheca, it widens along the hypotheca where it does not reach the . Red bodies often seen (Plate 1a-c). Resting cysts (not shown) are oval with a thick cell wall. Parvodinium cunningtonii is similar to P. elpatiewskyi in size and shape, and they tend to co-occur. See here our tips for distinguishing between these two species under the light microscope.
Ecology
Parvodinium cunningtonii is one of the species always present in the water column, contributing to total phytoplankton biomass up to 20-30%. The species was less abundant before the mid 1990s, when Peridinium gatunense bloomed every year, but has become more abundant with the decline of Peridinium gatunense in recent years (Fig. 1). Parvodinium cunningtonii tends to co-occur with Ps elpatiewskyi (Figs. 1, 2). Parvodinium cunningtonii abundance shows a typical annual cycle of population increase in March, peak in April and decline in May-June, with low abundances but continuous presence throughout the summer (Fig. 3). Cell size fluctuates seasonally, with largest size in winter, smallest in summer (Fig. 3). This feature is shared with several other Kinneret dinoflagellates.
Environmental conditions
Parvodinium cunningtonii cell abundance > 20 mL-1 occurred more frequently but not exclusively at Chloride concentration > 225 mg L-1, > 1050 μS cm-1, pH > 8.5, over the full range of (90-140 mg CaCO3 L-1, dissolved oxygen > 10 mg L-1, at a relatively narrow range of long-wave radiation (320-385 Watt m-2), but over the full range of short-wave radiation (100-350 Watt m-2) and water temperatures (14-30C), when the lake was stratified. The high levels of dissolved oxygen during its high abundance are unusual (Fig. 4). These conditions are extremely similar to those under which Parvodinium elpatiewskyi is abundant and indeed the two species nearly always co-occur (Figs 1,2).
Additional figures
Cite this record as: Dr. Tamar Zohary, Dr. Alla Alster. 16 June 2026. Electronic publication. Israel Oceanographic & Limnological Research. https://kinneret-algae-atlas.org/ Searched on —.
Further reading
- Hansen G, Flaim G. 2007. Dinoflagellates of the Trentino Province, Italy. J Limnol. 66:107-141.
- Pandeirada MS, Craveiro SC, Daugbjerg N, Moestrup Ø, Calado AJ. 2022. Ultrastructure and phylogeny of Parvodinium cunningtonii comb. nov. (syn. Peridiniopsis cunningtonii) and description of P. cunningtonii var. inerme var. nov. (Peridiniopsidaceae, Dinophyceae). Eur J Protistol. 86:125930.